

筆者:前藤 薫
•神を否定した寄生バチ•
「神によって世界は設計された」。これは近代以前の、キリスト教的世界観だ。その世界観では、地球上にこれほど多くの種類の生物が存在するようになったのは、「神による世界設計の証」とされていた。対して、近代科学は、事実と証明の積み重ねによって、上記の世界観を否定して宗教からの独立を果たした。「変異と自然選択こそ、多様な生物を生み出す原動力」と提唱し、キリスト教的世界観を進化論によって打ち砕いたのが、19世紀の生物学者ダーウィンである。
彼は植物学者グレイにあてた手紙のなかで、神による生命の創造を疑わざるを得ない事例として、寄生バチを取りあげている。毛虫を体内からむさぼり食べるヒメバチのような冷酷無情な生物を、慈悲深い全能の神が、創造されるはずがない(つまり神は生物を設計していない)と述べている。ダーウィンが寄生バチの生態に並々ならぬ関心を抱いていたことが分かる。
•透明な翼をもつ昆虫の誕生•
本書の主役であるハチの仲間は、透きとおった膜状の翅をもつことから「膜翅目(Hymenoptera)」と呼ばれる。約15万の種(全昆虫の約16%)を擁する巨大な生物群であり、その3/4を寄生バチと狩りバチが占める。哺乳類の種数は約6000種、鳥類が約9000種なので、桁違いの種多様性をもつことがお分かりいただけるだろう。
昆虫には30以上の目(order)と呼ばれる系統群が知られるが、膜翅目を含む「5大目」はとりわけ種数が多く、大繁栄している。これらのうち半翅目(セミ、カメムシ、アブラムシ)を除く、膜翅目、鞘翅目(コウチュウ)、双翅目(ハエ、カ、アブ)、鱗翅目(チョウ、ガ)の4目は、他の生物にはあまり見られない珍しい特徴をもっている。それは、幼虫と成虫の姿や生態が大きく異なる完全変態(holometabolism)という生活スタイルである。完全変態昆虫は、トンボやバッタ、カメムシなどの不完全変態昆虫から進化した新しいタイプの昆虫だが、植物が大森林を形成した石炭紀の中頃(約3億5000万年前)にはすでに出現していたようである。
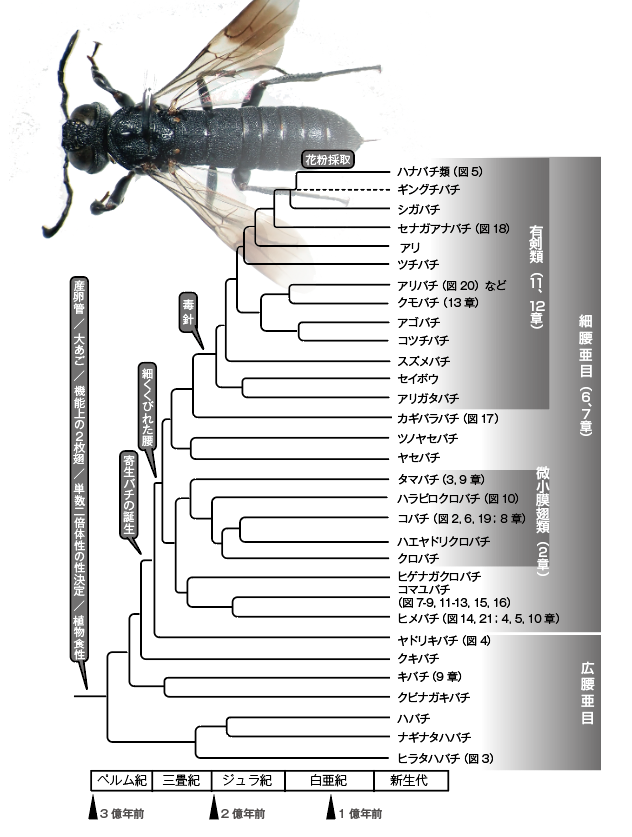
図1.膜翅目昆虫の系統発生。多数の遺伝子の発現解析に基づいて系統関係を推測し、化石によって年代を補正してある。各系統の階級名(上科、科)は省略し、複数系統を含む分類群は破線で示した。Peters et al. (2017) をもとに描く。
なかでも膜翅目は、最も早くに他の完全変態昆虫と袂を分かち、独自の道筋をたどって進化している。最も古いハチの化石は約2億2600万年前(三畳紀)のものだが、遺伝子系統解析の結果から約2億8100万年前(ペルム紀)には現在みられる多様なグループへの進化がすでに始まっていたと推定されている(図1)。 膜翅目は、他の昆虫にはあまり見られない「新しい特徴」と、多くの昆虫が退化させてしまった「原始的な特徴」をあわせ持っている(表1、図1)。

表1.膜翅目昆虫の主な特徴
では、これらの中で膜翅目の繁栄にとりわけ深く関わっている「道具」は何れだろうか。本章ではとくに、産卵管(ovipositor)と大あご(大顎、大腮 mandible)に注目したい。いずれも原始的な器官でありながら、ハチの生活に深く関わっている。これらの機能や関連する生態を中心に紹介しながら、ハチの進化と繁栄の秘密に迫りたい。専門的な細かい話はなるべく排除したので、ぜひ最後までお付き合いいただきたい。
•ふたつの古い道具 産卵管と大あご•
ふたつの器官のうち産卵管は、文字どおり卵を産むために使われる管である。だが、ただの単純な筒ではない。昆虫の産卵管は腹部の先端にある2対の付属肢(手足のもとになる突起物)が長く伸びて組み合わさったものであり、それらをすり合わせるように動かして卵が送られる。産卵管には神経が走っており、卵を間違いなく産み込むための感覚器官がある。産卵管は生きた器官なのである。
スズムシやコオロギは土中に深く卵を産み込むために長い産卵管を用いており、セミやイトトンボは植物体に産卵するときに鋭くとがった産卵管を使っている。だが、多くの完全変態昆虫は(普段は邪魔になる)産卵管を放棄して、必要な時にだけ腹部の先端部を長く伸ばしてその代わりにしている。ところが、膜翅目は原始的な産卵管を温存して、植物や昆虫の表皮を貫いて産卵したり(図7、11)、獲物を素早く麻酔する(図18)のに利用している。
では、もうひとつの器官、大あごとはどんなものだろう。我々は上あごと下あごで上下にものを噛むが、昆虫は左右から噛み合わせる一対の大あごをもっている(図2、9)。昆虫の頭部の大部分は脳ではなく、大あごを動かすための筋肉によって占められている(動画01)。バッタやトンボを思い浮かべてもらうと分かるように、大あごはもともと食物を噛み砕いたり引き裂いたりするための器官である。しかし、ハチの大あごの使い方はかなり異なる。食事のためではなく、羽化場所から外界に脱出する通路を開けたり、獲物を巣に運んだりするために大あごを利用している。 産卵管や大あごが、実際にどのように使われているのか見てみよう。
•ベジタリアンから始めた•
さまざまな証拠から、原始的な膜翅目昆虫は植物だけを食べるベジタリアン(植物食者)であり、産卵管を使って植物組織に卵を産み込む習性をもっていたと考えられている。ヒラタハバチ(図3)やハバチの仲間は、鋸をひくように産卵管を使って植物組織に切れ目を入れて卵を産み込む。幼虫は植物の葉を食べて育つと地面に落下し、天敵の少ない土中に潜って蛹化する。やがて羽化した成虫は、大あごを使って土をかき分けながら地表に現れる。

卵塊を保護するオオアカズヒラタハバチ(Cephalcia isshikii )の母親。トウヒの針葉を鋸状の産卵管で切開し、そこに卵の柄を埋め込んで固定する。母親は大あごをかざして天敵から卵を守る。
クキバチやキバチの仲間の成虫は、錐をもむように長い産卵管を植物の茎や幹に刺し込んで産卵し、幼虫は植物の内部を食べて育つ。なかでもキバチの習性には注目すべきものがある。キバチは栄養分に乏しい木部を幼虫の餌として利用するために微生物(キノコ)と共生しており、母親は産卵するときに(体内にもっている)キノコの菌糸とその成長を促す分泌物を木の幹に注入する(第9章に詳しい)。やがて成熟した幼虫は木の中で蛹化し、成虫は大あごを使って硬い木部に孔を開けて脱出する。
このように産卵管と大あごは、ベジタリアンだった原始的なハチが植物に産卵し、外界に脱出するために無くてはならない器官であった。やがてハチが肉食者(動物食者)へと進化するなかで、これらはさらに重要な役割を担うことになる。
•たぶん植物のなかで起こった 寄生バチの誕生•
約2億5000万年前のペルム紀末期には地球規模の環境変動によって三葉虫などの古いタイプの生物が絶滅し、やがて恐竜や翼竜の時代を迎える。ちょうどそのころ、ハチの世界でもその運命を決定づける大進化があいついで起きている。「寄生バチの誕生」と「くびれた腰の獲得」である(図1)。 最初の事件は植物の茎(幹)のなかで起きたようだ。植物組織を食べていたハチの幼虫が、たまたま出くわした昆虫の幼虫に齧り付いたらしい(蛹化前の動かない幼虫だったのだろう)。そうしてみると昆虫の幼虫は栄養に富んでいるので、植物を食べ続けるよりもずっと速く大きくなれた。やがて母バチは、植物の内部にひそむ昆虫の幼虫を狙って産卵管を刺し込むように進化したに違いない。寄生バチ(parasitic wasps)の誕生である。
あなたがハチの卵だとしよう。茎のなかで動き回る昆虫の幼虫にそのまま産み付けられては、そのうちに圧殺されてしまい危険である。だが大丈夫、そうならないように、母バチは麻酔液を注入して昆虫の幼虫が動かないようにしておいてくれる。植物を食べるハチの産卵管の付け根には、卵を滑らかに送り出す潤滑物質や卵が安全に育つために植物組織に働きかける成分を作る分泌腺がある。寄生バチでは、それが変化して他の昆虫を麻痺させる毒液(venom)を作り出す毒腺になったようだ。 ヤドリキバチの仲間(図4)は、このようにして誕生したであろう原始的な寄生バチの面影を留めている、生きた化石のようなハチである。だが、その姿があまり「ハチらしく」見えないのは、なぜだろう。

図4.トサヤドリキバチ(Ophrynopus tosensis)。腹部に収納された長い産卵管を繰り出して材内に生息する昆虫の幼虫に産卵する、原始的な寄生バチである。写真提供:渡辺恭平。
•くびれた腰と「2枚翅」•
「ハチらしさ」とは何か、と問われてまず思いあたるのは、腰のくびれであろう。原始的な植物食性のハチは、胸部と腹部が幅広くつながっているので「広腰亜目(Symphyta)」と呼ばれ、あまりハチらしく見えない(図3)。最初の寄生バチであるヤドリキバチも、まだ広腰(つまり寸胴)のままである(図4)。しかし、寄生生活をおくるようになって間もなく、寄生バチは付け根が細くくびれて動かしやすい腹部を手に入れている(図6〜8、10、11、14、16~20)。動かしやすい腹部は、昆虫を狙ってピンポイントで産卵管を刺し込むのに好都合である。こうして、「細腰亜目(Apocrita)」と呼ばれる「ハチらしい」ハチが出現した(図1)。
くびれた腰の進化はハチの飛翔能力の向上とも関わりがある。上手に飛べる昆虫の代表格は双翅目(ハエ、アブ、カ)だが、膜翅目もそれに次ぐ飛翔の名人だ。両者には「2枚翅」という共通点がある。双翅目の翅はたしかに2枚だが、膜翅目は4枚の翅をもつ。しかし、膜翅目の前翅と後翅はスライド式の構造によって連結されており(図5)、機能上は完全な2枚翅である。鳥やコウモリもそうだが、翼は2枚のほうが操りやすい。
ハエやハチの翅は筋肉に直接付着しているのではなく、胸部のしなやかに変形する骨格構造に取り付けられている。胸部には多数の筋肉が前後・上下に走り(図6、動画02)、それらの収縮と弛緩によって骨格が変形することによって、微妙な翅の動きがコントロールされている。マッチ箱に2枚の翅を取り付けて、その外枠を変形させることで翅を動かす様子を想像していただければと思う。
だが、ハバチやキバチなどの広腰亜目では胸部がまだ幅広く腹部とつながっているために、胸部の変形が強く制約を受けている。これに対して細腰亜目では、胸部が腹部から分離されることによって胸部骨格の自由度が高まり、より巧みに飛翔することが可能になっている。その際、腹部の付け根の部分は胸部に取り込まれて一体化し、残された腹部との間に自在に動かせる関節構造が形成された。このようにして、細腰亜目のハチに特有の「付け根が細くくびれて動かしやすい腹部」が形づくられた。

図5.セイヨウミツバチ(Apis mellifera)の後翅の前縁に並ぶ翅鉤(hamuli)。前翅の後縁のレールにはまってスライドする構造になっている。写真提供:徳平拓朗。
胸部と腹部の大再編によって、飛翔による高い探索能力と産卵管の優れた操作性を同時に手に入れた寄生バチの生活を、詳しく見てゆこう。
•動きを封じて寄生する 殺傷寄生•
寄生バチは、真の寄生者(parasite)ではない。真の寄生者は植物や動物から栄養分を横取りしても、それらを積極的に殺すことはしない。しかし、寄生バチは寄生した寄主(host、宿主とも呼ばれる)をほぼ必ず殺してしまう。この性質は捕食者(predator)に近いので、寄生バチは捕食寄生者(parasitoid)と呼ばれる。映画『エイリアン』に登場する地球外生命体(ヒトに捕食寄生する)を思い浮かべてもらっても良いが、もちろん寄生バチがヒトに寄生することはない。
寄生バチは最後には寄主を殺してしまうが、その過程によってふたつのグループに分けられる。ひとつは産卵時に寄主を永久麻酔して、事実上殺してしまう「殺傷寄生者(idiobiont)」である。もうひとつは産卵後もしばらくの間は寄主に自由な行動と発育をゆるすので、「飼い殺し寄生者(koinobiont)」と呼ばれる。
原始的な寄生バチは、ヤドリキバチ(図4)がそうであるように、植物体内にひそむ昆虫の幼虫に寄生する殺傷寄生者であったと考えられている。このタイプの寄生バチは、今でも広く見られる。彼らは産卵する前に産卵管を使って寄主に毒液を注入し、動きを封じてから、その体表に卵を産み付ける。毒液には麻酔成分だけでなく、寄主が死んでも腐らないように腐敗を防ぐ物質が含まれている。孵化した幼虫は短期間のうちに、麻痺した寄主の体液をすすって成長する。寄生する対象の幼虫や蛹は、生きた植物の茎や捲き葉、枯れ木の樹皮下、あるいは卵や繭のなかに隠れているので、寄生バチは外側から長い産卵管を刺し込んで毒液の注入と産卵をおこなう(図7)。寄生される幼虫や蛹はシェルターに守られているので、寄生バチの卵や幼虫も天敵に襲われる危険は少ない。

図7.サツマイモの茎にひそむイモゾウムシの幼虫に産卵するイリムサーキバラコマユバチ(Bracon yasudai)。
写真提供:上里卓己。
産卵管を含めると世界最長クラスの昆虫であるウマノオバチ(Euurobracon yokahamae)(図8)も、長い産卵管を駆使して木の中にひそむカミキリムシの幼虫あるいは蛹に産卵する殺傷寄生者である。このハチが長い産卵管をどのように使って卵を産み付けるのか長らく謎であったが、最近の研究によって、木の幹の表面から直接産卵管を刺し込むのではなく、幹の割れ目からカミキリムシの孔道に侵入して、孔道のなかで何回も向きを変えながら最終的に産卵管の先端を寄主に到達させることが明らかになった。だが、首尾よく寄生に成功したあと、木の中で羽化した新成虫がどうやって奥深い孔道から脱出するのか少し心配になる。そこで役に立つのが大あごである(図9)。ウマノオバチの丈夫な大あごは、孔道につまった木くずや樹皮を砕いて脱出孔を開けるために使われる。

図8.ウマノオバチ(馬尾蜂)。産卵管を含めた体長は約20cm に達する。描画素材提供:伊藤誠人
ところで、寄生バチや狩りバチの幼虫はウンコをしないことをご存じだろうか。彼らの肛門は閉じており、蛹になるとそれが開いて繭の中に宿便を排出する。幼虫が便をしない習性は、寄主の腐敗を防いで寄生を成功させるのに役立っているのだろう。

図9.ウマノオバチの顔。丸い大きな頭部には、分厚い大あごを動かすための筋肉が詰まっている。写真提供:渡辺恭平。
•卵の殻は安全なシェルター•
寄生バチが進化した三畳紀からジュラ紀にかけて(約2億5000万~1億4500万年前)は、他の完全変態昆虫が勢いを増した時代であった。植物の茎にひそむコウチュウやガの幼虫は、長い産卵管と丈夫な大あごを備えた殺傷寄生バチにとって、まさに格好の餌資源であったに違いない。だが、安全に寄生するための保護シェルターは植物組織でなくても良い。卵殻や繭に守られた昆虫の幼虫も絶好のターゲットである。
寄生バチは小型化しながら、先を争うように昆虫の卵に寄生する習性を進化させた(第2章に詳しい)。その結果、そもそも小さな昆虫であるチャタテムシやアザミウマの卵に寄生するコバチ類は、体長0.2mm以下という究極の小型化を達成している。ちなみに単細胞生物であるゾウリムシの大きさがちょうど0.2mm程度である(もちろん寄生バチは多細胞生物である)。卵寄生バチは、バッタやカマキリ、ナナフシ、さらにはトンボやアメンボ(図10)など、あらゆる昆虫の卵に寄生している。水中を泳いで産卵したり、寄主の成虫にしがみ付いて産卵場所まで便乗したりと、小さな体をフルに活かしている。

図10.アメンボタマゴクロバチ(Tiphodytes gerriphagus)。体長約1mm。水中を飛ぶように泳いでアメンボ類の卵に産卵する。Nishimori et al. (2018)より転載。
•育てながら寄生する 飼い殺し寄生•
だが、殺傷寄生には致命的な弱点がある。麻酔された寄主は天敵に対して無防備なので、天敵にとってみれば、何の努力もなしに魅力的なご馳走にあずかれるようなものである。そのため、殺傷寄生バチが利用する寄主は植物組織や卵殻、繭などのシェルターによって守られているものにほぼ限られており、葉っぱの上で自由に動き回る昆虫やクモには寄生できない。白亜紀以降(約1億4500万年前~)になると自由に歩きながら葉っぱを食べるチョウやガが繁栄を始めるが、そうした幼虫を利用できないのは残念だ。さて、どうしたものか。
名案がある。開けた場所で活動する寄主に安全に寄生するには、寄主を麻酔せずに、そのまま体内にひそんで寄生すれば良いではないか。そうすれば、アリやクモなどの天敵は、動くことのできる寄主が自分で追い払ってくれる。体内でゆっくりと成長すれば良いので、卵のサイズを小さくして、たくさんの卵を産むこともできる。
寄主に自由な行動と発育を許したまま寄生する寄生バチ、すなわち飼い殺し寄生バチは、殺傷寄生バチから何度も独立に進化している。その多くは寄主の体内に寄生する内部寄生者だが、体表に取りつく外部寄生者もいる。飼い殺し寄生バチは、チョウやガの幼虫はもちろん、アブラムシやカメムシ、クモ、さらにはアリやコウチュウの成虫(図11)、まだ稀ではあるがチャタテムシやバッタにいたるまで、自由に歩き回るさまざまな寄主を利用して繁栄している。

図11.ヤマナラシハムシの成虫に産卵するハラボソコマユバチの一種(Microctonus sp.)。ハムシの頭部と胸部の間の柔らかい膜質部を狙って正確に産卵管を刺し込んでいる。
もっとも、飼い殺し寄生を成功させるには、ふたつの難問を解決しなければならない。まず、活発に動き回る寄主に接近し、安全に産卵するための高い認知能力と運動能力が必要である。長期間の麻酔は許されないので、反撃のすきを与えずに素早く産卵する必要がある(プロローグ、図11)。チョウやガの幼虫はそれほど機敏ではないので大いに利用されているが、アリやクモを利用できる飼い殺し寄生バチが少ないのはそのためだろう。それでも挑戦は続いており、第4章にはアリヤドリバチの生態が、第5章にはクモと格闘するクモヒメバチの姿が描かれている。
ふたつ目に、昆虫の体内に寄生するにはその免疫系を無力化する必要がある。昆虫も強力な免疫システムをもっているので、何も対策をとらなければ寄生バチの卵や幼虫は病原体と同じように排除されてしまう。寄主の体内環境を整えるために寄生バチが繰り出すさまざまな妙技は、第6、8章に詳しく紹介されている。
•魔の時間が進化させた多様性•
メリットにはリスクがつきものである。飼い殺し寄生の利点は開けた場所で活動する寄主に安全に寄生できることだが、最後になって大きな落とし穴が待っている。開けた場所ではアリやクモのような天敵が、寄主の体から脱出する寄生バチの幼虫を待ち構えているのである。アリに食べられない丈夫な繭を作り終えるには一昼夜を要するので、無防備な幼虫はこの間を耐え忍ばなければならない。繭を完成させた後も、羽化するまでは危険な状態が続く。こうした「魔の時間」を、寄生バチはどのようにして凌いでいるのだろうか。 チョウやガの幼虫に寄生する飼い殺し寄生バチには、寄主幼虫が成熟するまで体内に留まって、寄主が繭を作り終えてからその中に脱出するものが多い。確かに安全なのだが、時間がかかる。そこで、プロローグでご紹介したギンケハラボソコマユバチのように糸を吐いてぶら下がって繭を作るものや、寄主の皮膚を硬化させてマミーの内部で蛹化するものもある(図12)。だが、もっと積極的な方法で身を守るものがいる。
皆さんなら、一世一代の人生イベントに妨害者がやってくる恐れがある場合、どうされるだろうか。ボディーガードを雇うのも一案であろう。寄生バチのなかには、直前まで自分が寄生していた昆虫をボデーカードに仕立てて、妨害者から身を守るものがいる。たとえば、チョウやガの幼虫に寄生するサムライコマユバチの仲間は、脱出して繭を作った後もしばらくは寄主幼虫を生かして天敵を追い払わせる(図13)。テントウハラボソコマユバチ(Dinocampus coccinellae)も寄主したテントウムシの成虫を操って繭を守らせるが、こうした寄主操作には寄生バチに共生していてテントウムシの神経系に感染するRNAウィルスが関与しているという。また、クモに寄生するクモヒメバチ類は、蛹化直前になるとクモの行動を変化させて、安全に繭を作るのに適した形状の網を張らせる(第5章に詳しい)。何とも巧みではないか。

図12.カモドキバチの一種(Aleiodes sp.)と寄主幼虫のマミー。寄主幼虫を硬化させて、そのなかで蛹化する。

図13.ブランコサムライコマユバチ(Glyptapanteles liparidis)の集合繭とマイマイガの幼虫。寄主のマイマイガ幼虫はコマユバチが脱出したあとも繭の近くに留まって捕食者をふり払う。
•闇夜への進出と「アメバチ風貌」•
寄生バチの多くは昼間に活動する。しかし、ガ類の幼虫の多くが天敵を避けて夜間に活動するので、それらの寄生バチには夕暮れどきから夜間にかけて飛翔するものが少なくない。夜中にライトを灯すとたくさんの寄生バチが集まるが(第14章に詳しい)、いちばん多いのは体全体が黄色いために「アメバチ(飴色のハチ)」と呼ばれるヒメバチの仲間である(図14)。
アメバチ以外にも、プロローグで紹介したハラボソコマユバチやカモドキバチ(図12)の仲間も夜行性である。そして奇妙なことに、それらは系統が遠く離れているにもかかわらず、どれも共通して「黄色い体」に「長い触角」と「大きな単眼(頭部の背面にある3個の眼)」をもっている。
寄生バチになったつもりで、夕暮れ時の薄暗い森の様子を想像してみてほしい。大きな単眼があれば、わずかな月明かりを頼りにして飛び回ることができる。長い触角は、暗闇のなかでにおいを手がかりにして寄主を探すのに役立つに違いない。また、夜間であれば黒い色素によって有害な紫外線から身を守る必要がないし、警告色や隠蔽色による擬態(後ほど詳しく説明する)も無意味なので、体色は一様に黄色でよいだろう。
この「アメバチ風貌(ophionoid facies)」と呼ばれる特異な姿かたちは、異なるグループの寄生バチがそれぞれ同じように闇夜に適応した収斂進化の結果であろう。

図14.サキグロホシアメバチ(Enicospilus ramidulus)。ヤガ類の幼虫に寄生する。夕暮れから夜間にかけて活動する寄生バチの多くが、黄色い体に長い触角と大きな単眼をもっている。写真提供:清水 壮。
•小さな寄主を大きく育てる•
飼い殺し寄生の利点は、開けた場所でくらす昆虫やクモに安全に寄生できることだけではない。寄主が卵のうちに産卵することによって、やがて幼虫が植物組織に深く潜り込んで産卵が難しくなる昆虫にも寄生できる。卵-幼虫寄生(egg-larval parasitism)と呼ばれる妙技である。
たとえば、ヨシノメバエコマユバチ属(Polemochartus)は初夏、水際に繁茂するヨシの新芽に産まれたばかりのヨシノメバエ類の卵に産卵する。孵化したハチの幼虫は、ハエの幼虫がヨシの茎に潜って大きく育つまでじっと待っていて、ハエが成熟するころになってその体内で成長を始める。翌春、ヨシの茎のなかで羽化したハチの成虫は、「外向きに咬む」ユニークな大あご(図15)を使ってヨシの茎に孔を開けて脱出する。
どんぐりを放置しておくと、白くて丸っこい幼虫がはい出てくることをご存じだろうか。シギゾウムシの仲間の幼虫である。シギゾウムシの成虫はどんぐりがまだ柔らかなうちに、長い口吻を使って産卵するための孔を開け、そこに卵を産み込む。幼虫は堅くなったどんぐりの内部を食べて育つので、寄生バチが幼虫に産卵することは難しい。そこでコマユバチの一種(Triaspis curculiovorus)(図16)は、ゾウムシの産卵孔に長い産卵管を挿入して、産卵直後のゾウムシの卵に卵を産み付ける。寄生されたゾウムシの幼虫は、健全な幼虫と同じようにどんぐりから脱出すると土に潜って冬を越すが、当然、翌年になって羽化してくるのはコマユバチの成虫である。どんぐりの殻は堅く、土に潜った幼虫に産卵することもできないので、産卵直後の卵は寄生バチがシギゾウムシに寄生できるほぼ唯一のチャンスである。
卵-幼虫寄生は、丈夫な産卵管をもたない小さな寄生バチが大きな寄主を利用するにも都合がよい。トビコバチの仲間は短い産卵管しかもたないので大きなガの厚い皮膚をもつ幼虫には産卵できないが、卵になら産卵できる。卵に産み込まれたハチの胚子は寄主の体内で多数に増殖し、寄主幼虫が成熟するころになると大量のハチがそれを食い破って羽化する(第8章に詳しい)。
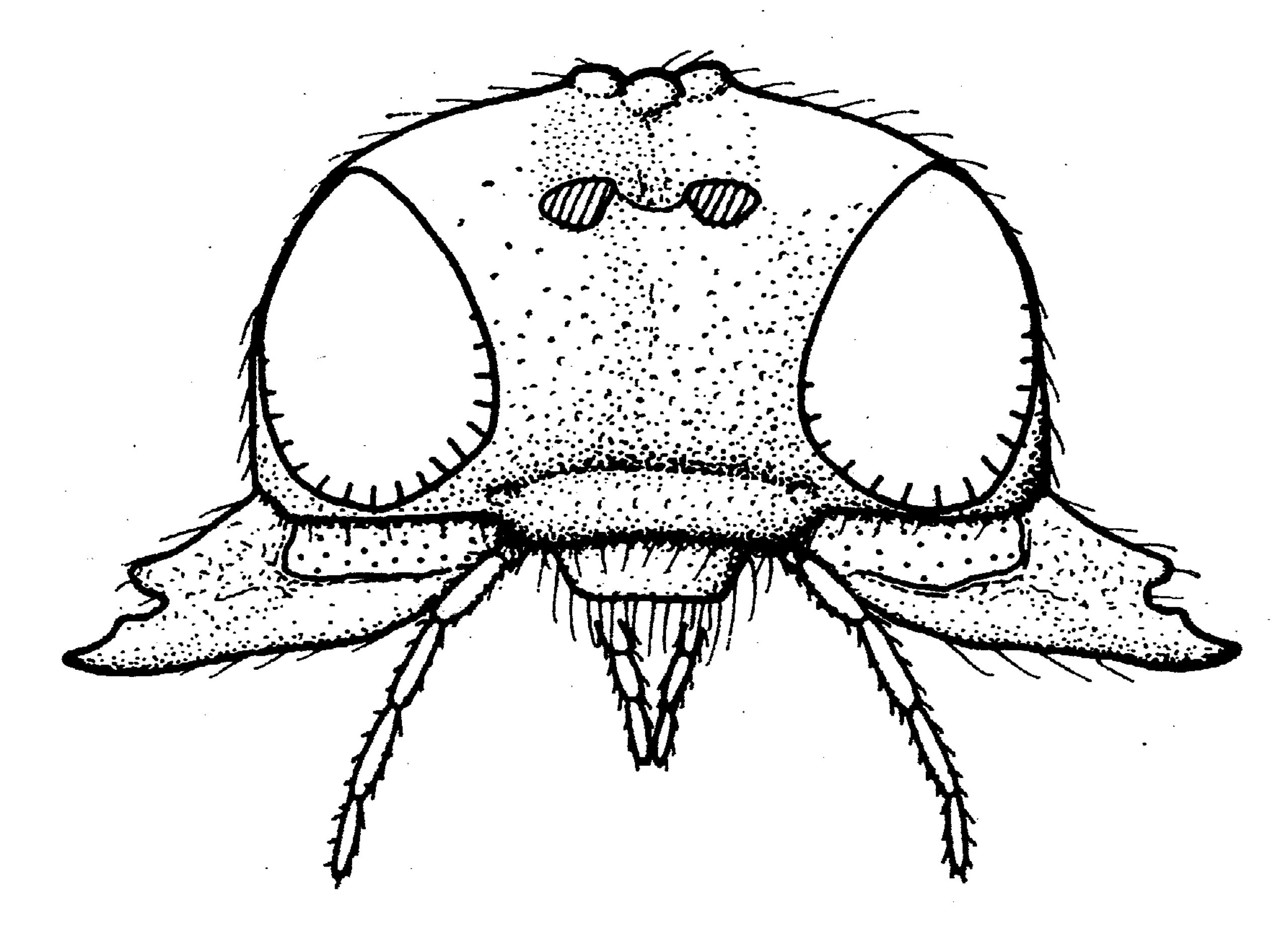
図15.ニホンヨシノメバエコマユバチ(P. nipponensis)の頭部。外向きに大きく開く大あごは、ヨシの茎を破って脱出するときに使われる。Maeto (1983)より改写。

図16.シギゾウムシ類に寄生するコマユバチの一種(T. curculiovorus)。長い産卵管をゾウムシの産卵孔に挿入してその卵に産卵する。
•においや動きを手がかりにして探す•
食事するお店を選ぶとき、美味しそうなにおいのするところに入るように、多くの寄生バチはまず植物のにおいを手がかりにして寄主に近づく。そのあと、植物の食べあとにのこされた唾液や糞のにおいから寄主を特定して産卵する。昆虫に食べられている植物や昆虫に共生する微生物からは独特のにおいが発生することがあるので、そうしたにおい成分を手がかりにするものもあり、第9章に詳しく紹介されている。
お店の見た目や店がまえも重要である。ハムシの成虫に産卵するハラボソコマユバチは、視覚をたよりにして動く寄主に近づくと、頭部と胸部の間の柔らかい部分(あるいは肛門)を狙って正確に産卵管を挿入する(図11)。だが、寄生バチの視覚認識に関する研究はまだ少ない。 美味しいお店は覚えておきたい。寄生バチも、刺激に単純に反応するだけでなく、さまざまな情報を学習することが知られている。たとえば、タバコガ類の幼虫に産卵するサムライコマユバチの一種(Microplitis croceipes)は、触角で寄主の糞に触って、糞に残る植物の欠片から発生するにおい成分を学習する。その後は、その植物から発生するにおい成分を手がかりにして、遠くからでも寄主を探すことができる。
卵-幼虫寄生バチは、卵に産卵する際に幼虫の糞のにおいのような確かな寄主情報を使えないので、より慎重に産卵をおこなう。たとえば、ハマキガ類に寄生するハマキコウラコマユバチ(Ascogaster reticulatus)は、植物成分を手がかりにして寄主の卵塊に近づくと、そこに残された鱗粉のにおいを嗅いでから卵に産卵管を挿入する。だが、すぐに産卵するのではなく、卵内に存在する物質の成分(アミノ酸)を確かめてから卵を産む。寄主の識別には、触角はもちろん、産卵管にある感覚器も大活躍である。
•寄生バチにも寄生する•
寄生バチはあらゆる昆虫に寄生するが、その寄生対象には寄生バチ自身も含まれている。普通は産卵管を用いて寄生バチの幼虫や蛹に産卵するが(動画03)、カギバラバチの仲間(図17)のやり方はかなり変わっている。
カギバラバチは数千個から一万個もの微小な卵を葉っぱの縁に産み付ける。卵はほとんどが無駄になるが、ごく一部は葉を食べたガやハバチの幼虫に飲み込まれて体内に入る。その幼虫がたまたま寄生バチ(あるいは寄生バエ)に寄生されたら、今度はその寄生バチ(寄生バエ)の幼虫に寄生して成育していくが、そうでなければ死んでしまう。

図17.キスジセアカカギバラバチ(Taeniogonalos fasciata)。産卵管は短く、腹部の先端が強く湾曲しているので「鉤腹蜂」と呼ばれる。写真提供:渡辺恭平。
さらに大きなご馳走にあずかろうと、ガやハバチの幼虫がスズメバチに捕獲されるのを待って、スズメバチの幼虫に寄生するものもいるが、宝くじに当たるようなものである。莫大な数の卵は、きわめて低い寄生成功率と釣り合っていることになる。どうして最初に卵を飲み込んだガやハバチの幼虫に寄生しないのか分からないが、植物を食べる昆虫の体には植物に由来するさまざまな毒物質が含まれているので、より安全な食べ物であろう寄生バチや寄生バエの幼虫と出会うのを待って寄生するのかも知れない。
•巣に運び込む寄生バチ 狩りバチの誕生•
寄生バチの話題がひと段落したので、いよいよ狩りバチ(hunting wasps)についてお話ししたい。もっとも、狩りバチの幼虫の食性は、永久麻酔されて動かない寄主(獲物)の体液をすする殺傷寄生者と変わらない。では殺傷寄生バチと狩りバチの違いは、どこにあるのだろうか。
ことの起こりはジュラ紀の初め、スズメバチなどを含む有剣類(Aculeata)と呼ばれる系統群の出現にさかのぼる(図1)。有剣類の産卵管は文字どおり麻酔専用の剣(毒針)に変形していて、卵は普通、産卵管を通らずに腹部先端から直に産み落とされる。原始的な有剣類は殺傷寄生バチそのものだが、そこから狩りバチが何回も独立に進化し、さらに狩りバチの一部はハナバチへと進化している(図1)。
殺傷寄生バチが麻酔した寄主は、自分の身を守ることができない。そのため原始的な殺傷寄生バチは、はじめから植物の茎や捲き葉などのシェルターに隠れている寄主を探して産卵する。だが、犬が餌を横取りされないように穴を掘って隠すように、捕らえた寄主(獲物)を保護シェルターに運び込んでもよいだろう。狩りバチは、まさにこの方法を採用した。
強力な毒針で麻酔した獲物を、シェルターである「巣」に運び込んで幼虫に食べさせるというユニークなやり方によって、狩りバチは餌メニューをいっきに増やした。チョウやガの幼虫はもちろん、ゴキブリ(図18)、バッタ、コオロギ、ハムシ、ハエ、クモなど、自由に動き回るあらゆる節足動物が狩りの対象になっている。
クモ類は豊富に存在する餌資源だが、危険な動物なのでうまく利用できる寄生バチや狩りバチは少ない。クモバチの仲間は勇敢にもクモを捕らえて巣に運び込む習性を進化させて大成功している(第13章に詳しい)。他方でクモの幼体に外部寄生するクモヒメバチの仲間も、クモの行動を巧妙に操作するなど特異なやり方でクモを利用している(第5章に詳しい)。前者は狩りバチとして、後者は飼い殺し寄生バチとして、全く異なる技を駆使しながらクモを攻略する両者を比較していただきたい。
狩りバチの巣の構造や材料はさまざまである。植物の茎のような自然の孔を借用するもの、土や枯れ木に孔を掘るもの、さらには土や小石、植物繊維などを集めて巣を作るものもいる。初めは獲物を巣に運んでから卵を産み付けていたのだろうが、巣に卵を産んでから狩りに出かけるものも現れた。単独で巣を作っていたものが、集団で営巣するようになると家族や社会ができる(第13章に詳しい)。自分では巣作りや餌集めをしないで、近縁種の巣に侵入して餌ごと乗っ取ってしまう連中(労働寄生者 cleptoparasite)も現れた。巣はダニなどの共生生物の棲み家にもなるので、そこにはさらに複雑で豊かな生態系が形成されることになる。詳しくは第12章をご一読いただきたい。
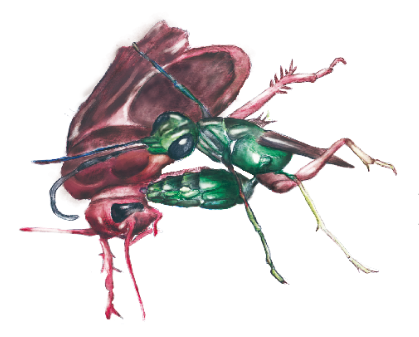
図18.ゴキブリを攻撃するエメラルドゴキブリバチ。最初の一撃で前肢を麻痺させた後、脳を狙って2回目の刺撃を行う。
Moore et al. (2018)より作画。
•ゾンビにされるゴキブリ•
寄生バチから狩りバチへと進化する過程で変化したのは、産卵管だけではない。もうひとつの古い道具である、大あごの役割も大きく変わった。寄生バチの大あごは、成虫が外界に脱出する通路を開けるときに使われる。いわばトンネル掘削の専用ツールである(後述するように、雄同士が闘ったり、雄が雌を運んだりするときに使われることもあるが珍しい)。しかし、狩りバチでは、通路を開けるだけでなく、獲物を捕獲して運ぶ、巣作りのため土や木を掘る、巣材を運ぶなど、人間の手のように万能な器官へと変化しており、まさに大活躍である。
だが、大あごでは運べないほど大きな獲物を狩るハチもいる。エメラルドゴキブリバチ(Ampulex compressa)は、毒針による1回目の突き刺し(刺撃)でゴキブリの幼虫の前肢を麻痺させた後、ゴキブリに催眠術をかけるかのように、2回目の刺撃によって脳に毒液を注入してその行動を支配する(図18)。ゴキブリはハチに触角を引っ張られると素直に誘導されて、自らの肢で歩いてハチの巣に向かう。自らの墓穴へと歩む姿は、まさにゾンビ(生ける屍)に他ならない。こうした巧妙な行動操作には、神経伝達物質であるドーパミンのほか、毒液に含まれる複数の特異なペプチド類(アミノ酸の短い結合体)が関わっているという。この研究は北米でおこなわれたものだが、近縁のセナガアナバチ類は日本にも生息しており、同じようにゴキブリを狩っているので観察していただきたい。
•父親をもたない息子たち•
膜翅目の特徴は性の仕組みにもある。我々の性別は性染色体の組み合わせ(XXなら雌、XYなら雄)によって決まるが、ハチの性別は単数二倍体性(あるいは単倍数性 haplodiploidy)と呼ばれる仕組みによる。このシステムでは、1セットの染色体をもつ個体(単数体)は雄に、2セットもつ個体(二倍体)は雌に育つ。雌は減数分裂をへて単数体の卵子をつくり、雄はそのまま単数体の精子をつくる。単数体の卵がそのまま産まれると雄に、精子と一緒に産まれると受精して二倍体の雌に育つ。お気づきのとおり、なんとも風変わりなことに、雄には父親がいないのである。
この特異な性決定システムには、どのような利点があるのだろうか? まず、母親は交尾しなくても息子を産める。さらに、交尾した雌は精子を体内に貯めておき、産卵する時に精子を卵にかければ娘、かけなければ息子というように、子の性を産み分けることができる。殺傷寄生バチの子の体サイズは産卵時の寄主の大きさ(資源量)によって決まってしまうので、大きな寄主には娘、小さな寄主には息子を産むものが多い。また、ひとつの寄主から多数の子が羽化してそこで交尾する寄生バチでは、息子同士が交尾相手(娘)を求めて無駄な競争(局所的配偶者競争 local mate competition)をしないように、息子を少なく産むことも知られている。性の産み分けができるのは何かと便利である。
それだけではない。有害な突然変異の多くは二倍体では発現しにくい性質(潜性、劣性)をもつので集団内に蓄積しやすい。しかし、ハチの雄は単数体なので潜性の有害遺伝子が発現して早々に集団から除去される。そのため、膜翅目では集団が小さくなっても近親交配による悪影響(近交弱勢 inbreeding depression)が現れにくく、新しい種の分化が進みやすいのではないかと考えられている。さらに、次節で述べるように、単数二倍体性の生物では真社会性が進化しやすい。
もっとも、これらの「利点」はどれも単数二倍体性の「結果」であって、それを進化させた「要因」とは考えにくい。ハチの性システムの進化については、第7章の議論をご堪能いただきたい。
•真社会性のパラドックス•
アリやミツバチがつくる真社会性(eusociality)と呼ばれる社会システムは、我々の社会とどこが違うのだろうか。我々の社会ではさまざまな役割を分担している個人が、それぞれ子を産み育てることができる。しかし、真社会性の社会では生殖も分業化されていて、自分では子を産まずにもっぱら他者の生殖を助ける個体が存在する。膜翅目では少なくとも3回、不妊の姉(働きバチ)が母親(女王)を助けて、妹(新女王)を養育するタイプの真社会性が進化している。だが、自分では子を残さない性質の遺伝子が自然選択によって集団内に広がることなど、あり得るのだろうか?
このパラドックスに答えたのは、イギリスの進化生態学者ハミルトンである。彼は膜翅目の性決定システム(単数二倍体性)に着目した。そして、ハチの姉妹は染色体を1セットしかもたない父親から全く同じ染色体を受け継ぐため、姉妹間の遺伝的な近さ(3/4)が母娘間のそれ(1/2)よりも大きくなることを指摘した。そうすると、どうだろう。自分の遺伝子を残すときに、子を介するよりも妹を介した方が、効率が良くなる。そのため膜翅目では、姉が子を産まずに、母親を助けて妹を育てるタイプの真社会性が進化したのだと説明されている(ハミルトンの3/4仮説)。
しかし、それだけではハバチや寄生バチではなく、狩りバチにおいて真社会性が何回も進化したことを理解できない。狩りバチと寄生バチの違いを思い起こしてほしい。狩りバチだけが子を育てるための「巣」をもち、そこで家族生活を始めている。母親世代と娘世代が同居して助け合える条件が整うことによって、はじめて真社会性が進化したと考えられる。狩りバチにおける社会性の進化については、第13章もお読みいただきたい。
他方で寄生バチには、女王の存在しない、全く異なるタイプの真社会性が見られる。1個の卵から発生した遺伝的に等しい幼虫のなかに、自分を犠牲にして兄弟姉妹を守る兵隊カーストが生じるのである。第8章には、トビコバチ科の寄生バチにおける、こうしたカーストの分化や性比調節の仕組みが詳しく紹介されている。
•生物農薬とバンカープラント法•
感染症を治療するときに抗生物質の使い方を間違えると耐性菌が出現してしまう。同じように、害虫に対しても漫然と同じ殺虫剤を使用していると抵抗性(耐性)をもつ害虫が進化してしまう。また、ミツバチなどの花粉媒介昆虫を利用する際にも殺虫剤は使えない。そこで、殺虫剤だけでなく色々な方法を組み合わせて害虫を防除する研究が進められており、寄生バチや狩りバチも害虫を退治するための防除素材(「生物農薬」と呼ばれることもある)として大いに期待されている。
たとえば、アブラムシ類に寄生するギフアブラバチ(Aphidius gifuensis)は野菜類の害虫アブラムシの防除に用いられているが、ここでちょっとした工夫をご紹介したい。天敵アブラバチを菜園に放す前に、野菜以外の植物(ムギ類)を植えて無害なアブラムシ類(天敵アブラバチの寄主になる)を放しておくのである。そうすると、そこであらかじめ天敵アブラバチを増やしておけるので、害虫アブラムシに対する防除効果を高めることができる。この方法は、「植物銀行」に天敵を預けて増やすような仕組みなので、バンカープラント法(banker plant system)と呼ばれる。
寄生バチを天敵として利用する事例は各章にも紹介されているが、詳しくは仲井まどからの『バイオロジカル・コントロール』を紐解いてほしい。
•ベジタリアンへの回帰•
膜翅目は食べ物を植物から動物に変えて多様に進化したが、後になってベジタリアンに回帰したものも多い。幼虫に与えるタンパク源を肉から花粉に変更した狩りバチの仲間は、ハナバチ(花蜂bees)として繁栄している(図1)。また、第3章に紹介されているタマバチの仲間のほか、コバチやヒメバチの仲間の寄生バチにも植物に虫こぶ(ゴール)を作る習性がしばしば進化している。もともと植物を食べていたハバチ類にも虫こぶを形成するものが多いので、ハチは植物組織を刺激して肥大化させる能力を早くからもっていて、それを繰り返し利用しているのかも知れない。
イチジク類の花粉媒介者(ポリネーターpollinator)であるイチジクコバチの仲間も、昆虫食からベジタリアンに回帰した元寄生バチである(図19)。熱帯林には果実をつける植物が多いが、たいていはある時期に集中して実をつける。そのため、果実食の鳥類や哺乳類は、いつも果実をつけているイチジク類に大いに依存している。それらの動物はイチジク類を食べて命をつなぎながら、多種類の樹木の種子を散布する役割を担っている。そこで、イチジク類は熱帯林生態系の維持に欠かせない重要な生物、すなわち「キーストーン生物(keystone organism)」とされる。
それでは何故、イチジク類はいつも果実(花)をつけているのだろうか。それは、イチジク類がイチジクコバチ類だけに花粉媒介を頼っているからであろう。イチジクコバチ類はもっぱらイチジク類の雌花を食べて育ち、しかも成虫は短命である。専属の花粉媒介者を飼いつなぐため、イチジク類は絶え間なく花を咲かせる必要がある。

図19.イヌビワコバチ(Blastophaga nipponica)の雄(左)と雌(右)。交尾はイヌビワの花嚢の中で行われるので、雄バチ(左)は翅をもたず、雌をめぐって闘うのに適した鎧のようなつくりの体をしている。写真提供:松尾和典。
•翅を捨てるのはどちら?•
何かを捨てることも進化である。昆虫学の授業では、翅の獲得こそ昆虫を繁栄に導いたもっとも重要な要因だと教わる。しかし、アリは翅を捨てることでより高度な社会性を進化させたし、アリ以外の膜翅目でも翅の欠失や退化がしばしば起きている。
先ほど紹介したイチジクコバチ類の多くは、イチジク類の花嚢の中で交尾をすますので雄バチは翅をもたないことが多い(図19)。だが、ハチの全体を見渡すと翅を失うのは雌バチの方が多いようだ。
インドネシアの熱帯林では、ペルガハバチの一種(Cladomacra taoi)の翅をもたない母バチが、枯れ木の樹皮の下で卵塊を見守る様子が観察されている。幼虫は知られていないが、樹皮下でカビを食べているようだ。
昆虫の繭やクモの卵嚢に寄生するハネナシヒメバチ属(Gelis)や古い畳を食べるタバコシバンムシに寄生するシバンムシアリガタバチ(Cephalonomia gallicola)など、徘徊しながら寄主を探索する寄生バチの雌もしばしば翅を失っている。有剣類の巣に潜って幼虫や蛹に産卵するアリバチ科の雌も翅をもたないが(アリに似ているのでそう呼ばれる)、雄は雌よりも大きくて翅をもつものが多い(図20)。面白いことに雄は交尾したまま、あるいは大あごでくわえて、無翅の雌を産卵場所まで運ぶ。大あごの新しい使い方である。

図20.アリバチの一種(Pherotilla oceanica)(左)とルイスヒトホシアリバチ(Smicromyrme lewisi)(右)の雄(有翅)と雌(無翅)。雄は交尾したまま(左)、あるいは大あごでくわえて(右)、翅のない雌を運ぶ。写真提供:岡安樹璃也。
•なりすまし 守るか攻めるか•
スズメバチに刺されたことのある方なら、黒と黄色の縞模様をみるとその時の痛みの記憶がよみがえるので、もうハチの巣には近づくまいと思われるだろう。黒と黄色の縞模様はスズメバチが身を守るのに役立っている。身を守る擬態(mimicry)には、警告色を用いる標識的擬態と保護色を用いる隠蔽的擬態がある。警告色は自分が危険なものだと天敵に思わせて、攻撃を思いとどまらせる仕掛けである。ふたつのタイプがあって、危険な動物同士が同じような警告色をまとって天敵の学習効果を強化していればミュラー型擬態、無害な動物が危険な動物に似せていればベイツ型擬態というように区別される。
黒と黄色の縞模様は、スズメバチやアシナガバチなど、毒針をもつ狩りバチに共通した警告色なのでミュラー型擬態と考えられる。同じような縞模様を、ハナアブやハバチなど、か弱い昆虫が真似ているのはベイツ型擬態ということになる。ところが、ヒメバチの仲間など大型の寄生バチにもよく似た縞模様をまとうものがいる(図21)。ヒメバチは毒針を持たないのでベイツ型擬態かと思われるが、触ると産卵管で刺すようにもがき、刺されるとチクリと痛いので、ミュラー型擬態として機能している可能性も否定できない。

図21.キオビコシブトヒメバチ(Metopius browni)。黒と黄色の縞模様は警告色であろう。写真提供:渡辺恭平。
つぎに隠蔽的擬態の例を見てみよう。エダナナフシは体幹や肢を木の枝に似せて背景に溶け込んでいるが、身を隠す背景は視覚的なものばかりではない。アリの巣に同居する昆虫たちは、働きアリに攻撃されないように、アリと同じにおい成分(ワックス)をまとって化学的に擬態している。ケアリ類の巣に同居するエイコアブラバチ(Paralipsis eikoae)もそうした好蟻性昆虫の仲間である。アリの巣に飛来すると、最初に働きアリに体を擦りつけてワックスを奪う。こうしてアリに化けたアブラバチは、アリから口移しで餌をもらい、アリの目を盗んでアリが保護しているナシマルアブラムシに近づいて産卵する。アブラバチの化学的擬態は、ただ身を守るためだけでなく、繁殖のためにアリを操作する攻めの戦術といえる。
アリの巣に住みついたアブラバチの産卵管は短く、大あごは小さい。しかし、アブラムシの体内に産卵し、その硬いマミーから脱出するには、それらが無くてはならない。ふたつの古い道具は脈々と受け継がれ、ハチたちの個性的な生活を支え続けている。
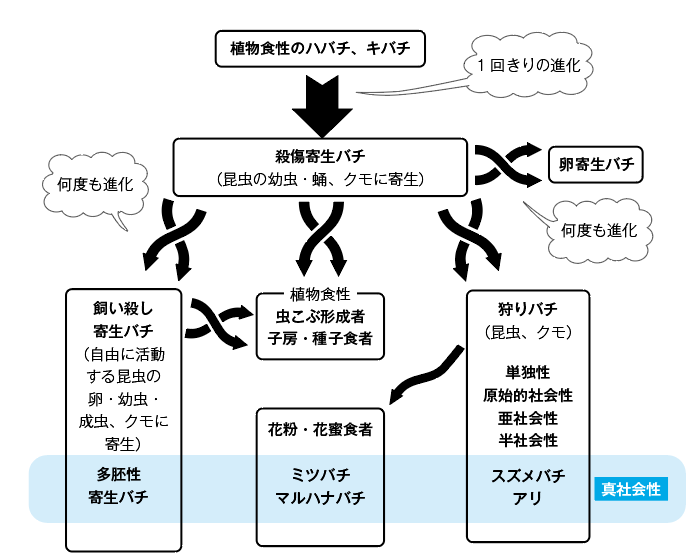
図22.膜翅目における大進化。植物食から殺傷寄生への進化は1回きりだが、他の進化は何回も独立に起こっている。
•ハチの世界の大進化•
本章の締めくくりに、膜翅目がたどった大進化の道筋をまとめてみたい(図22)。ベジタリアンとして起源したハチの祖先から、昆虫を麻酔して食べる殺傷寄生バチへの進化は1回きりだったと考えられる。その後、殺傷寄生バチからは卵寄生バチや狩りバチが何度も進化した。また、寄主の活動性を保ったまま寄生する飼い殺し寄生バチへの進化も繰り返し起こっていて、彼らは寄主の体内環境や行動を操作する術を身につけて新たな餌資源を開拓しつつある。一方で寄生バチや狩りバチは何回もベジタリアンに回帰し、虫こぶ形成者や種子食者、花粉媒介者に進化した。ひんぱんに真社会性が進化しているのも膜翅目の際立った特徴である。
この章で注目した産卵管と大あごは、完全変態昆虫の多くが放棄してしまった古い道具だが、ハチはそれらをうまく使いこなして独特の風変わりな世界を繰り広げている。以下の各章では、その豊かな世界をさらに深くご堪能いただきたい。
貴重な画像をご提供いただいた伊藤誠人、松尾和典、西森敬晃、小倉一輝、岡安樹璃也、清水 壮、徳平拓朗、上里卓己、渡辺恭平、山岸健三の各氏およびマイクロフォーカスX線CT(図2、6)の撮影にご協力くださった株式会社島津製作所の櫛引敬嗣氏に厚くお礼申し上げる。
主な参考文献ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
日高敏隆監修(1998)『日本動物大百科第10巻昆虫Ⅲ』平凡社.[生態写真が素晴らしい]
平嶋義宏編(2017)『図説 日本の珍虫 世界の珍虫:その魅惑的な多様性』北隆館.
岩田久二雄(1971)『本能の進化 : 蜂の比較習性学的研究』眞野書店.[復刊版(1981)サイエティスト社][狩りバチ研究の古典的名著]
加賀玲子ら(2018)「ウマノオバチEuurobracon yokahamae (Dalla Torre, 1898) (Insecta: Hymenoptera: Braconidae) の生活史,特にその寄主について」
『神奈川県立博物館研究報告(自然科学)』47, 59-66.
太田泉編(2016)『ギフアブラバチ利用技術マニュアル』農研機構.
丸山宗利ら(2013)『アリの巣の生きもの図鑑』東海大学出版会.
松本吏樹郎ら(2012)『ハチまるごと!図鑑』大阪市立自然史博物館. 百瀬邦泰(1998)「熱帯林の生物多様性をなぜ、どのようにして保全するのか」『地球環境』3, 21–27.[イチ
ジクコバチの花粉媒介者としての重要性が紹介されている]
仲井まどから編(2009)『バイオロジカル・コントロール:害虫管理と天敵の生物学』朝倉書店.[寄生バチの生理と行動、利用について詳しい]
坂上昭一(1970)『ミツバチのたどったみち:進化の比較社会学』思索社.[新装版(2005)新思索社]
佐藤芳文(1988)『寄生バチの世界』東海大学出版会.[日本語で書かれた最初の総説]
杉浦直人ら編(2002)『ハチとアリの自然史』北海道大学図書刊行会.
スコット・リチャード・ショー[藤原多伽夫訳](2016)『昆虫は最強の生物である』河出書房新社.[Shaw, S. R. (2014) Planet of the Bugs: Evolution and the
Rise of Insects. The University of Chicago Press][昆虫の進化史について分かりやすく紹介されている]
Darwin Correspondence Project, “Letter no. 2814,” accessed on 11 October 2019, https://www.darwinproject.ac.uk/letter/DCP-LETT-2814.xml
Dheilly, N.M. et al. (2015) Who is the puppet master? Replication of a parasitic wasp-associated virus correlates with host behaviour manipulation. Proc. R. Soc. B 282, 20142773.
Gauld, I.D. and Bolton, B. (ed.) (1988) The Hymenoptera. Oxford University Press, Oxford. [膜翅目の全貌が紹介されている。イラストや写真も多くて分かりやすい]
Kainoh, Y. and Tatsuki, S. (1987) Host egg kairomones essential for egg-larval parasitoid, Ascogaster reticulatus Watanabe (Hymenoptera: Braconidae). I. Internal and external kairomones. J. Chem. Ecol. 14, 1475-1484.
Lewis, W. and Takasu, K. (1990) Use of learned odours by a parasitic wasp in accordance with host and food needs. Nature 348, 635–636.
Maeto, K. (1983) A systematic study on the genus Polemochartus Schulz (Hymenoptera, Braconidae), parasitic on the genus Lipara Meigen (Diptera, Chloropidae). Kontyu 51, 412-425.
Moore, E.L. et al. (2018) Ampulexins: A new family of peptides in venom of the Emerald Jewel Wasp, Ampulex compressa. Biochemistry 57, 1907-1916.
Nishimori, T. et al. (2018) Morphological and genetic analyses suggest some strains under Tiphodytes gerriphagus (Hymenoptera: Platygastridae: Scelioninae), an egg parasitoid of water striders. Jpn. J. Sys. Entomol. 24, 277–282.
Peters, R.S. et al. (2017) Evolutionary history of the Hymenoptera. Curr. Biol. 27, 1013-1018.
Shinohara, A. (1986) A new apterous sawfly from Sulawesi, Indonesia (Hymenoptera: Pergidae: Perreyiinae), and the pleural origin of the ventral region of the sawfly mesothorax. Sys. Entomol. 11, 247-253.
Takada, H. and Hashimoto, Y. (1985) Association of the root aphid parasitoids Aclitus sappaphis and Paralipsis eikoae (Hymenoptera, Aphidiidae) with the aphid-attending ants Pheidole fervida and Lasius niger (Hymenoptera, Formicidae). Kontyu, 53, 150-160.
Quicke, D.L.J. (1997) Parasitic Wasps. Chapman & Hall, London. [寄生バチ生物学の本格的な入門書]
Quicke, D.L.J. (2009) Hymenoptera (ants, bees, wasps). In V. H. Resh and R. T. Carde (ed.) Encyclopedia of Insects, Second Edition. pp. 473-483. Academic Press, USA.[膜翅目の概要と重要文献が紹介されている]
Quicke, D.L.J. (2015) The Braconid and Ichneumonid Parasitoid Wasps: Biology, Systematics, Evolution and Ecology. Wiley-Blackwell, Oxford.[Quicke (1997)の続編でもある]
