
ここでは本の発売に先駆けて、「コラム① 絶滅した魚たち」をご覧になれます。ぜひご覧いただいて、本書を手に取る参考にしてください。
分子系統解析は、脊椎動物の出現が地質時代のカンブリア紀(5億4000万〜4億8000万年前)に相当する5億年前あたりであることを示唆している。カンブリア紀は、ひとつ前の地質年代のエディアカラ紀(6億2000万〜5億4000万年前)と比べて出土する動物化石がはるかに多く、その種類の豊富さや多様性からカンブリア紀初期に多細胞生物の多様化が急激に進んだとされている(カンブリア爆発)。化石記録によれば、カンブリア爆発は動物の歴史全体の1.7%程度の期間に起きており、この短い間に現生のほとんどの後生動物と同じボディプランを有する生物が出現したらしい。脊椎動物のいくつかの形態形質(体幹部と明確に区別できる頭部や眼、明瞭な鰓構造など)を備えた化石種(ミロクンミンギアMyllokunmingiaやハイコウイクチスHaikouichthysなど)も、このカンブリア紀の地層から見つかっている。最初期の脊椎動物については系統関係が不明瞭なので詳細は省くが、円口類と顎口類の分岐がかなり早い段階で起こったことはほぼ間違いない。円口類は顎がないために無顎類とも呼ばれるが、古い時代に絶滅してしまった脊椎動物化石種の中には円口類とは異なるグループ(甲皮類Ostracoderms)の無顎類が存在していた。現在では、この円口類以外の顎をもたない化石無顎類を顎口類に含める考えが広まりつつある(31)。「顎をもたない顎口類」というのは違和感があるが、化石無顎類は厳密にはステム顎口類という位置づけである。つまり、無顎類という単語は単系統ではなく、特定のグレード(単系統ではない便宜的分類群)を指すものである(詳しくは図10)。円口類以外の化石無顎類では甲皮や鱗など一部の骨格が顎口類と同じくミネラル化しており、また、対鰭や対の内部嗅覚器官など現生顎口類により近いボディプランを呈する種もいる。ちなみにヤツメウナギやヌタウナギも硬い歯をもつが、これはケラチンというタンパク質を主成分とした角質歯であり、象牙質をもつ石灰化した顎口類の真歯とは異なる。化石種を含む全ての円口類にミネラル化した骨格の証拠は存在しない。
上下の顎をもつ真の有顎脊椎動物の化石はシルル紀(4億4000万〜4億2000万年前)から見られるようになる。顎の獲得により高次捕食者となった顎口類は、淡水・海水両方の環境で繁栄した。初期の顎口類の4つの系統(板皮類、類、軟骨魚類、硬骨魚類)の化石の多くはデボン紀(4億2000万〜3億6000万年前)から見つかっており、デボン紀は魚の時代と呼ばれる。硬骨魚類と軟骨魚類が単系統であることについての異論はないが、初期の顎口類の起こりについては今もはっきりしておらず、特に下に示したように板皮類と棘魚類の系統位置はいまだに議論の的である。どちらも現在では単系統とみなされず、板皮類を顎口類のステムグループ、棘魚類は軟骨魚類のステムグループとして扱うことで落ち着いている。以下に4つの系統の特徴を紹介しよう。
棘魚類……微小な鱗を纏い、ほぼ全ての鰭の前端に棘を備えた棘魚類はオルドビス紀後期からペルム紀前期まで生き延びており、その最後の生き残りは有名なペルム紀の棘魚類アカントーデスAcanthodesである。棘魚類は淡水を中心に栄えたが、残念なことに古い年代の完全な頭蓋の化石がほとんど存在しなかったため、より後期のアカントーデスについての解析から、全ての棘魚類が硬骨魚類のステムグループだと結論づけられてきた。ところが、その後アカントーデスの頭蓋に軟骨魚類の特徴が見出され、またデボン紀初期の棘魚類であるPtomacanthusの頭骨が硬骨魚類よりも板皮類や軟骨魚類のそれと似た特徴を示すことが明らかにされた。よって近年では硬骨魚類よりも軟骨魚類の祖先種であったとする見方が強くなっている。さらに後述の板皮類の化石記録も、棘魚類を軟骨魚類のステムグループとする根拠のひとつとなっている。
板皮類……全長10mに達するダンクルオステウスが有名だが、他にも多様な形態を示す化石種が含まれる。全ての板皮類は化石無顎類に似た装甲で頭や体幹が包まれている。板皮類のいくつかの種は軟骨魚類と類似の外生殖器クラスパーをもち、体内受精であった。この特徴から板皮類を軟骨魚類の姉妹群とする説も唱えられたが、今では軟骨魚類のクラスパーとの相同性は疑問視されている。2010年代に化石板皮類のEntelognathusとQilinyuの発見によって、これまで硬骨魚類にしかないと考えられてきた顎骨要素をもつ板皮類の存在が明らかになり、板皮類は軟骨魚類のステムグループではないと考えられるようになった。つまり、板皮類は硬骨魚と軟骨魚の分岐より前の枝に存在したとされ、顎口類ステムグループとしての地位は確かなものとなった。板皮類には、棘魚類や軟骨魚類のように断片化した小さな皮歯がない事実もこの仮説を支持する。板皮類は当初は淡水に栄え、その後大部分が汽水や海水へと生息域を拡大した。
軟骨魚類……さて本章で扱ってきた軟骨魚類はというと、化石記録で有名なものは長い間最古の全身化石とされてきたデボン紀後期の軟骨魚類クラドセラケCladoselacheだろう。ただこの最古記録は2000年代に入っていくつかの化石によって更新されている。そのひとつDoliodusは100年以上もの間棘魚類だと考えられてきたが、その後に見つかった頭部を含む化石の歯板や頭蓋、そして肩帯や胸鰭の解析によって、棘魚類から初期軟骨魚類が生じた分岐付近に位置する種だと考えられている。化石軟骨魚類の生息域はほとんどが海水だが、デボン紀後期から三畳紀まで栄えたXenacanthida目の軟骨魚類は淡水棲であった。この事実は、軟骨魚類がいつ尿素調節能を備えるようになったかを知る上でヒントになるかもしれない。
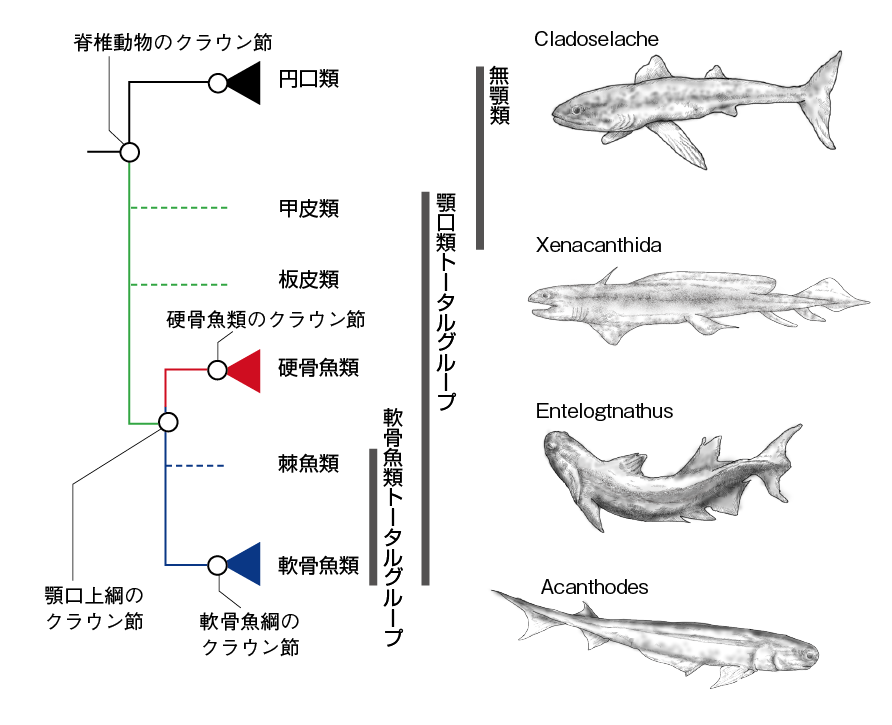
図10. 化石種を含めた系統樹。現生の脊椎動物については分子情報も含めて精度の高い分類が可能だが、化石種についてはトータル(全)、クラウン(冠)、ステム(幹)グループといった考えを採用する。トータルグループは枝分かれした後の種を全て含む分類で、クラウングループは特定のグループの最後の共通祖先とそこから枝分かれした全ての種の呼称である。ステムグループはトータルグループからクラウングループを除いた分類群。例えば左の図で棘魚類は、軟骨魚類のクラウングループには含まれない、ステム軟骨魚類である。(Cladoselache、Xenacanthida、Acanthodes:http://spinops.blogspot.com/より作図、Entelognathus:https://dx.doi.org/10.1371/journal.pone.0170929より作図)
硬骨魚類……最後に硬骨魚類だが、当然その化石は軟骨魚類よりもよく保存されている。ただ軟骨魚類と同様に古い化石が出てくるのは特定の地域に限られるため情報が乏しく、肉鰭類と条鰭類の分岐などの初期進化についての詳細はよくわかっていない。条鰭類の祖先の存在を示す確実な証拠が、肉鰭類よりもかなり遅いデボン紀中期からしか発見されないことは謎のひとつである。一方で肉鰭類とわかる化石はシルル紀後期から見つかっており、デボン紀初期に肉鰭類は既にシーラカンスとDipnotetrapodomorpha (ハイギョと四肢動物:Rhipidistia)に分かれていた。Rhipidistiaの祖先は海水あるいは汽水環境に生息し、その後淡水へと生息域を移していったらしい。そして今からおよそ3.5億年以上も前のデボン紀のうちに、化石両生類は陸上に進出していたのである。
化石や古代環境というのは人々の興味をひきつけてやまないが、上述の進化の物語は現時点で最ももっともらしいもののひとつを取り上げたに過ぎない。新しい化石の発見や解析技術の進歩によって定説はいく度となく覆されており、今も白熱した議論が繰り広げられている。
